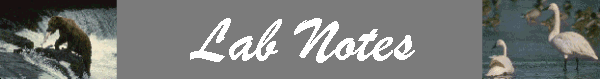
Review concepts of predator-prey interactions, including properties that affect stability.
Learn what factors can influence the shape of a functional response and a numerical response.
Predator: an organism that uses other live organisms (prey) as an energy source and, in doing so, removes the prey individuals from the population (Colinvaux 1986).
Parasitoids: any number of so-called parasitic insects whose larvae live within and consume their host, usually another insect, inevitably leading to the death of the host (Ricklefs 1990).
Parasite: an organism that consumes part of the blood or tissues of its host, usually without killing the host.
Herbivore: an organism that consumes living plants or their parts; herbivores may function either as predators (consuming whole plants) or parasites (consuming living tissues but not killing their victims).
How would the prey population grow in the absence of the predator?
How many prey does each predator consume, i.e., what is the functional response?
How does predator density change when prey density changes, i.e., what is the numerical response?
|
|
We might also ask about the stability of the predator-prey interactions. |
In studying predator-prey interactions it is important to distinguish between factors that influence predator abundance and factors that influence predator searching efficiency (i.e., a predator's ability to find and consume prey; Hassell 1978). This distinction has resulted in recognizing two predator responses: functional and numerical.
A functional response measures how many prey are needed to maintain a predator over a specific time period (e.g., number of moose killed/wolf/100days vs. moose density; Krebs 2001). Three general types of functional response are recognized. Each one illustrates a predator-prey interaction that is either stabilizing or destabilizing.
Type 1 Functional Response – as prey increases, predator search time decreases so there is an initial rapid increase in prey numbers taken. However, there usually is some upper limit to the number of prey a single predator can handle per unit of time. A type I response is neutral until an upper limit is reached, then it tends to be destabilizing.
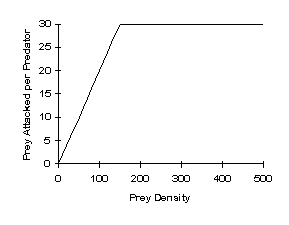
Type 2 Functional Response – similar response to Type 1, but the upper limit is reached more slowly. Tends to be destabilizing. This is distinguished from a Type 1 response because this accounts for prey handling time by the predator. The handling time will depend on: (1) time spent searching and killing prey, (2) time spent eating prey, and (3) time spent cleaning or resting after feeding (Holling 1959).
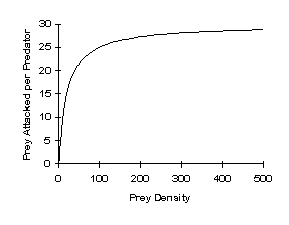
Type 3 Functional Response – sigmoid-shaped curve possibly due to prey switching, which can occur when the predator concentrates on the prey species with the largest densities. This causes a delay between when the prey begins to increase and when the predator detects the increase and switches effort. Tends to be stabilizing.
A numerical response measures how predator density increases through reproduction based on prey density (e.g., wolf density vs. moose density). It is important to realize predators do not search at random for their prey, but instead concentrate on patches of high prey density. The ability of predators to aggregate in patches of high prey density is critical in determining how effective a predatory can be in limiting prey populations (Krebs 2001).
Type 1 Numerical Response – linear relationship where predator's K is set by prey density. Neutral in terms of stabilizing effect.
Type 2 Numerical Response – increase in predator numbers with increased prey abundance, but predator population reaches a threshold where something else limits the population (e.g., territories, nest sites, etc.). Somewhat destabilizing.
Type 3 Numerical Response – a density-dependent predator response to prey abundance (sigmoid-shaped curve). Stabilizing effect up to some threshold level.
Type 4 Numerical Response – No predator response to changes in prey abundance; the number of predators per prey gets smaller. Destabilizing.
Type 5 Numerical Response – negative response in predator abundance occurs as prey population increases. Destabilizing.
|
|
You need to look at functional and numerical responses together in order to gain insight into the stability of predator-prey interactions. |
dH / dt = r H - b1 H P
where,
H = number of prey (H for herbivores) P = number of predators r = intrinsic rate of growth for prey population b1 = predation rate (coefficient expressing the efficiency of predation).
dP / dt = -m P + b2 H P
where,
b2 = rate of growth of the predator per unit contact with prey m = predator's intrinsic rate of growth (decline) in absence of prey
|
|
The simultaneous solution of the Lotka-Volterra equations for predation predicts that numbers of both predators and prey should oscillate, and the oscillations should be coupled (Coupled Oscillation Hypothesis). |
|
|
Numbers of both predators and prey circle a singular stable point in perpetually balanced imbalance, i.e., neutral limit cycle. The degree of oscillation about the equilibrium point is a function of initial population sizes. Perturbations may eventually result in extinction of prey and predator. |
Unrestricted exponential growth of the prey population in the absence of predators.
Environment is homogeneous.
Every prey has an equal probability of being attacked, e.g., no age-based selection.
Predators have an unlimited capacity for increase, i.e., the response of predators to prey is linear (i.e., a Type 1 numerical response).
Prey density has no effect on the probability of being eaten, i.e., a Type 1 functional response.
Predator density has no effect on the probability of a predator capturing prey.
No time lags.
 |
 |
 |
Revised: 06 December 2004