| Life tables are used to describe age-specific mortality and survival rates for a population. When this information is combined with fecundity data, life-tables can be used to estimate rates of population change (e.g., r, lambda, and R0). |
Cohort or age-specific or dynamic life tables: data are collected by following a cohort throughout its life. This is rarely possible with natural populations of animals. Note: a cohort is a group of individuals all born during the same time interval.
Static or time-specific life tables: age-distribution data are collected from a cross-section of the population at one particular time or during a short segment of time, such as through mortality data. Resulting age-specific data are treated as if a cohort was followed through time (i.e., the number of animals alive in age class x must be less than alive in age class x-1). Because of variation caused by small samples, data-smoothing techniques may be required (see Caughley 1977).
Composite - data are gathered over a number of years and generations using cohort or time-specific techniques. This method allows the natural variability in rates of survival to be monitored and assessed (Begon and Mortimer 1986).
Semelparity - Individuals that have only a single, distinct period of reproductive output in their lives, prior to which they have largely ceased to grow, during which they invest little or nothing in survival to future reproductive events and after which they therefore die (Begon et al. 1996:147). For annual species, this results in nonoverlapping generations. Examples other than annual plants and some insects?
Iteroparity - Individuals that normally experience several or many such reproductive events. During each period of reproductive activity the individual continues to invest in future survival and possibly growth. It, therefore, has a reasonable chance of surviving to reproduce again (Begon et al. 1996:147). Results in overlapping generations.
Birth pulse - reproductive activity is restricted to a specific breeding season. Begon et al. (1996) refer to this as "overlapping semelparity."
Birth flow - reproductive events merge into a single extended period.
| x = | age, measured in years or some other conventional unit. With longer-lived animals and plants this is often 1 year, but for voles it might be 1 week and for some insects 1 day. Often expressed as an interval, e.g., 0-1 years old. |
| nx = nx-1 - dx-1 = | the number of individuals surviving at the start of age interval x. Note: n0 = sum(dx) if dx expressed as numbers dying and the survival schedule is complete for all members of the cohort. |
| dx = nx - nx+1 = | the number of individuals of a cohort dying during the age interval x to x+1. Note: sometimes calculated as proportion dying. |
| qx = dx / nx = | finite rate of mortality during the age interval x to x
+ 1. Note: this parameter is least affected by bias in the sample and gives the most direct projection of the mortality pattern in a population. It is often used to make comparisons within and between species. |
| Sx = (nx-dx)/nx or 1 - qx = | finite rate of survival during the age interval x to x + 1. This parameter is used in harvest calculations and in population modeling. Note: finite rates cannot be added to get total survival rate for a longer period of time; however, finite survival rates are multiplicative (i.e., survival from age 0 to 3 = S0 x S1 x S2). |
| lx = nx / n0 = | the proportion (scaled from 0 to 1) of individuals surviving at the start of age interval x. You will also see lx expressed per 1,000, i.e., = (nx / n0)1000 . This parameter is used to plot survivorship curves (see comments below). |
| kx = log10 nx - log10 nx+1 = | killing power or a standardized measure of the intensity (rate) of mortality. Unlike qx-values, kx-values can be added to determine the mortality rate for a number of age classes. Note: because kx-values are calculated using log10, you must take the antilog of the common log to convert back to finite survival (i.e., S = 10-k). |
| mx = | fecundity rate (i.e., average number of female offspring produced per female in the population over some period of time, generally a year). |
| lx mx = | The mean number of female offspring produced by females in an age class. This information is used to calculate net reproductive rate (R0) and the instantaneous rate of change ( r ). |
| R0 = [sum (lx mx)] / l0 = | the net reproductive rate per generation. Or in other words, the mean number of female offspring produced by a female during her lifetime (i.e., the replacement rate). Note: R0 < 1 indicates the members of the population are not replacing themselves (i.e., the population is declining). R0 > 1 denotes an increasing population. R0 = 1 indicates a stationary or "stable" population. |
| G = Sum(lx mx x) / R0 = | the mean length of a generation. Other definitions include (1) the time elapsing between birth of a female and mean time (age) of birth of her offspring and (2) the average age that adult females give birth. |
| the finite rate of population change or the net reproductive rate over some time interval, which is a year in many cases. Lambda >1 indicates an increasing population, lambda=1 indicates a stationary population, and lambda<1 indicates a decreasing population. The use of this parameter is geared toward organisms that reproduce during a short breeding season (i.e., discrete growth or birth-pulse fertility). Note: your textbook uses "R" to denote lambda. | |
| r ~ = ln(R0)/G = ln(lambda) = | intrinsic or instantaneous rate of increase (i.e., the change in population size per individual per unit of time). An r>0 indicates an increasing population, r=0 indicates a stationary pop'n, and an r<0 indicates a declining population. Note: the equations listed only give you an approximation of r. You need to solve Euler's equation for a precise estimate of r (see Begon et al. 1996:165). |
| ex = Tx / Lx
= where Lx = (lx + lx+1) / 2 |
mean expectation of further life for individuals alive at the start of age interval x. The mean expectation of further life can be used as one way of compressing an entire life table into one number. It has limited application for wildlife studies, but is commonly used in the insurance business. |
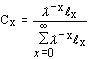 |
stable age distribution (i.e., where the proportion of the population in each age class remains constant over time). This is only achieved if the observed survival and fecundity schedules remain constant over a long period of time. Note: population projections from simple deterministic growth models (e.g., the exponential and logistic growth models) are based on the assumption that the population has a stable age distribution. |
NOTE: Population ecologists do not all use the same life-table notation. For example, Begon et al. (1996) use different symbols to denote the finite rate of increase and mean generation length. Nevertheless, data and calculations in the respective columns have the same meaning.
Age at Death Recorded Directly - The number of individuals dying in successive intervals of time is recorded for a group of individuals born at the same time. This is the most precise type of data available because it is based on a single cohort followed through time. The observed data are the dx column of the life table.
Cohort Size Recorded Directly - The number of individuals alive in successive intervals of time is recorded for a cohort of individuals. These data are similar to those obtained with Method 1, except that those surviving are tallied, not those dying. These data are also precise and specific to the cohort studied. Observed data are entered into the nx column of the life table.
Age at Death Recorded for Several Cohorts - Individuals are marked at birth and their death recorded, as in Method 1, but several cohorts are pooled from different years or seasons. These data are usually treated as if the individuals were members of one cohort and the analysis of Method 1 is applied.
Age Structure Recorded Directly - The number of individuals aged x in a population is compared with the number of these that died before reaching age x+1. The number of deaths in that age interval, divided by the number alive at the start of the age interval, gives an estimate of qx directly.
Ages at Death Recorded with a Stable Age Distribution and Known Rate of Increase - Often it is possible to find skulls or other remains that give the age at death of an individual. These data can be tallied into a frequency distribution of deaths and thus give dx directly.
Age Distribution Recorded for a Population with a Stable Age Distribution and Known Rate of Increase - In this case, the age distribution is measured directly by sampling. The number of individuals born is calculated from fertility rates.
Note: Methods 5 and 6 are based on the critical assumption that the rate of population change is known (or the population is stationary, i.e., r=0) and the age distribution is stable. Although methods 5 and 6 appear more realistic in terms of data collection, there are numerous ways in which time-specific life tables have been calculated incorrectly, e.g., from hunter kills (see Caughley 1977).
Caughley (1977) recommended that the cohort consist of at least 150 individuals when basing survival estimates on the stable-age assumption. However, age determination is difficult for many species, and if age is not measured carefully the resulting life table may be very inaccurate.
The main value of a life table lies in what it tells us about the population's strategy for survival, i.e., life tables help us to understand the dynamics of populations. For example, time-specific life tables, although often not meeting the assumptions necessary to estimate survival rates, are valuable to a manager of exploited populations because they show the existence of strong year classes or help identify weak age classes.
Although we have used age-structured schedules, for some organisms age is not the best life history variable on which to develop analyses of population change. For example, the stage of development (egg, larval, pupal, adult stages) of some insects may be a more important life history variable. If predation on fish is size-dependent, then size rather than age would be the appropriate stage-variable.
Opportunities to follow cohorts for long periods of time are rare, which precludes cohort analysis. Furthermore, the critical assumption of a stable-age distribution is so difficult to meet, that it limits the use of time-specific life tables (Johnson 1994).
Although life tables are rarely used in fisheries work, information on age or size composition of the population can be used to estimate survival (see Ricker 1975). Catch curves are probably the most common method for estimating mortality of exploited fish populations (see Ricker 1975, Krebs 1989).
Survivorship curves are usually created by plotting lx on the y-axis and age on the x-axis. Occasionally you may see nx plotted on the y-axis. The y-axis is usually logarithmic, i.e., log10 (lx), to allow comparisons among different studies and species. In other words, log transformations standardize the survivorship curve.
Mortality curves are created by plotting qx or kx against age.
Compare your survivorship/mortality curves to those described in Begon and Mortimer (1996:153-154) and Johnson (1992:433).
Begon, M., J. L. Harper and C. R. Townsend. 1996. Ecology: individuals, populations, and communities. 3rd edition. Blackwell Scientific Ltd., Cambridge, Massachusetts, USA.
Begon, M., and M. Mortimer. 1986. Population ecology: a unified study of animals and plants. 2nd edition. Blackwell Scientific Publications, Boston, Massachusetts, USA.
Caughley, G. 1977. Analysis of vertebrate populations. John Wiley & Sons, New York, New York, USA.
Johnson, D. 1994. Population analysis. Pages 419- 444 in T. Bookhout, editor. Research and management techniques for wildlife and habitats. Fifth edition. The Wildlife Society, Bethesda, Maryland, USA.
Krebs, C. J. 1989. Ecological methodology. Harper & Row Publications, New York, New York, USA.
Ricker, W. E. 1975. Computation and interpretation of biological statistics of fish populations. Fish Research Board of Canada Bulletin 191.
Revised: 10 November 2010